
In contrast to the linear chromosomes found in eukaryotic cells, the strains of bacteria initially studied were found to have single, covalently closed, circular chromosomes.1 The circularity of the bacterial chromosome was elegantly demonstrated by electron microscopy in both Gram negative bacteria (such as Escherichia coli) and Gram positive bacteria (such as Bacillus subtilis). Bacterial plasmids were also shown to be circular. In fact, the experiments were so beautiful and the evidence was so convincing that the idea that bacterial chromosomes are circular and eukaryotic chromosomes are linear was quickly accepted as a definitive distinction between prokaryotic and eukaryotic cells. However, like most other distinctions between prokaryotic and eukaryotic cells2, it is now clear that this dichotomy is incorrect. Not all bacteria have a single circular chromosome: some bacteria have multiple circular chromosomes, and many bacteria have linear chromosomes and linear plasmids.
Experimental evidence for multiple chromosomes and linear chromosomes initially came from studies using pulsed field gel electrophoresis (PFGE), an approach that uses alternating electric fields to separate large DNA molecules on an agarose gel. Subsequently genome sequencing projects have added to the list of bacteria with multiple or linear chromosomes.
Experimental evidence for multiple chromosomes and linear chromosomes initially came from studies using pulsed field gel electrophoresis (PFGE), an approach that uses alternating electric fields to separate large DNA molecules on an agarose gel. Subsequently genome sequencing projects have added to the list of bacteria with multiple or linear chromosomes.

The first convincing evidence that some bacteria have multiple chromosomes came from studies on Rhodobacter sphaeroides. Both molecular (Suwanto and Kaplan, 1989) and genetic (Suwanto and Kaplan, 1992) studies clearly demonstrated that R. sphaeroides has two large circular chromosomes. One of the chromosomes is 3.0 Mb and the other is 0.9 Mb. Genes encoding rRNAs and tRNAs required for translation, and metabolic enzymes are distributed between the two chromosomes. Multiple chromosomes have also been found in many other bacteria, including Agrobacterium tumefaciens, Rhizobium, Brucella, Paracoccus denitrificans, Ochrobactrum anthropi, Leptospira interrogans, Burkholderia, Vibrio cholerae, Deinococcus radiodurans, and many others from diverse groups of bacteria.
Furthermore, some bacteria have linear chromosomes. Borrelia3 have linear chromosomes and most strains contain both linear and circular plasmids; most of the bacteria in the genus Streptomyces4 have linear chromosomes and plasmids and some have circular plasmids as well. In addition, in some cases there may be a dynamic equlibrium between linear and circular forms of a DNA molecule. There is some evidence that linearization may be due to integration of a linear phage genome into the circular DNA molecule (Volff and Altenbuchner, 2000).
Linear chromosomes and plasmids were not discovered in bacteria until relatively recently. The first published evidence for linear chromosomes was in 1979, but because the techniques used at that time were limited5 and because the dogma that all bacterial chromosomes are circular was so entrenched, few people believed that linear chromosomes and plasmids occured in bacteria until 1989. By that time pulsed field gel electrophoresis had been developed, and this new technique provided convincing evidence that the chromosome of Borrelia burgdoferi was linear.
The ends of linear DNA molecules (called telomeres) pose two problems that do not apply to circular DNA molecules. First, since free double-stranded DNA ends are very sensitive to degradation by intracellular nucleases, there must be a mechanism to protect the ends. Second, the ends of linear DNA molecules must have a special mechanism for DNA replication.6 These problems are solved by features of the telomeres. Two different types of telomeres have been observed in bacteria: hairpin telomeres and invertron telomeres.
Furthermore, some bacteria have linear chromosomes. Borrelia3 have linear chromosomes and most strains contain both linear and circular plasmids; most of the bacteria in the genus Streptomyces4 have linear chromosomes and plasmids and some have circular plasmids as well. In addition, in some cases there may be a dynamic equlibrium between linear and circular forms of a DNA molecule. There is some evidence that linearization may be due to integration of a linear phage genome into the circular DNA molecule (Volff and Altenbuchner, 2000).
Linear chromosomes and plasmids were not discovered in bacteria until relatively recently. The first published evidence for linear chromosomes was in 1979, but because the techniques used at that time were limited5 and because the dogma that all bacterial chromosomes are circular was so entrenched, few people believed that linear chromosomes and plasmids occured in bacteria until 1989. By that time pulsed field gel electrophoresis had been developed, and this new technique provided convincing evidence that the chromosome of Borrelia burgdoferi was linear.
The ends of linear DNA molecules (called telomeres) pose two problems that do not apply to circular DNA molecules. First, since free double-stranded DNA ends are very sensitive to degradation by intracellular nucleases, there must be a mechanism to protect the ends. Second, the ends of linear DNA molecules must have a special mechanism for DNA replication.6 These problems are solved by features of the telomeres. Two different types of telomeres have been observed in bacteria: hairpin telomeres and invertron telomeres.
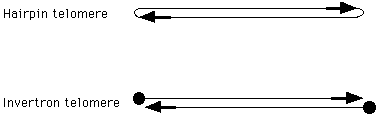
There are examples of linear DNA molecules in bacteria that are protected by both types of telomeres: palindromic hairpin loops are protected by the lack of free double-stranded ends, and invertron telomeres are protected by proteins that bind to the 5'-ends. Both of these mechanisms are also used by some phage, eukaryotic viruses, and eukaryotic plasmids.8
The two types of telomeres also solve the problem of DNA replication differently. Invertron telomeres have a protein covalently attached to the 5' ends of the DNA molecule (called the 5'-terminal protein or TP for short). DNA polymerase interacts with the TP at the telomere and catalyzes the formation of a covalent bond between the TP and a dNTP. The dNTP bound to the TP has a free 3'-OH group which acts as the primer for chain elongation. Replication of hairpin telomeres is less well understood. Apparently multiple hairpin sequences can pair to form concatemers that are replication intermediates.7
The important take-home point is that we are just beginning to appreciate the similarity of many processes once thought to be completely different between bacteria and eukaryotes, partly because we now have better tools for studying these processes and partly because most of the earlier studies focused on relatively few types of bacteria. The more we study a wider diversity of bacteria, phages, and plasmids, the more obvious it becomes that E. coli is an excellent model for dissecting broad features of molecular and cell biology, but not all bacteria do everything the same way. Furthermore, we have only recently begun to attack the molecular genetics of the Archae, and what we have learned so far suggests that this diverse group of prokaryotes share even more common features with the eukaryotes.
The circular genomes of mitochondrial and chloroplast are a notable exception to the rule that eukaryotic chromosomes are linear. However, this nicely fit into the dichotomy that eukaryotic chromosomes are linear and bacterial chromosomes are circular because these organelles seem to have evolved from entrapped bacteria.
Other examples include the presence of introns, and poly-A tails on mRNA.
This genus includes B. burgdorferi, the causative agent of Lyme disease.
Streptomyces make a wide variety of useful antibiotics, including streptomycin.
For example, linear DNA was precipitated in the most commonly used procedures for purifying bacterial plasmids, and the procedures for purifying chromosomal DNA relied upon the differential binding of ethidium bromide to "sheared DNA fragments" compared to circular DNA.
It is not intuitively obvious how the ends of a linear DNA molecule could be completely replicated. All known DNA polymerases require a pre-existing primer for initiation of DNA replication. The primer is usually a short RNA molecule with a free 3'-OH group that can be extended by DNA polymerase. If a linear DNA molecule was primed at one end, DNA synthesis could continue to the other end. However, once the primer is removed, the DNA corresponding to the primer could not be replicated.
The telomers at the end of chromosomes of most eukaryotic cells are replicated by a different mechanism: most telomeres are short GC-rich repeats that are added in a 5' to 3' direction by the enzyme telomerase (see Blackburn. 1991. Telomeres. Trends Biochem. Sci. 16: 378-381).
The two types of telomeres also solve the problem of DNA replication differently. Invertron telomeres have a protein covalently attached to the 5' ends of the DNA molecule (called the 5'-terminal protein or TP for short). DNA polymerase interacts with the TP at the telomere and catalyzes the formation of a covalent bond between the TP and a dNTP. The dNTP bound to the TP has a free 3'-OH group which acts as the primer for chain elongation. Replication of hairpin telomeres is less well understood. Apparently multiple hairpin sequences can pair to form concatemers that are replication intermediates.7
The important take-home point is that we are just beginning to appreciate the similarity of many processes once thought to be completely different between bacteria and eukaryotes, partly because we now have better tools for studying these processes and partly because most of the earlier studies focused on relatively few types of bacteria. The more we study a wider diversity of bacteria, phages, and plasmids, the more obvious it becomes that E. coli is an excellent model for dissecting broad features of molecular and cell biology, but not all bacteria do everything the same way. Furthermore, we have only recently begun to attack the molecular genetics of the Archae, and what we have learned so far suggests that this diverse group of prokaryotes share even more common features with the eukaryotes.
The circular genomes of mitochondrial and chloroplast are a notable exception to the rule that eukaryotic chromosomes are linear. However, this nicely fit into the dichotomy that eukaryotic chromosomes are linear and bacterial chromosomes are circular because these organelles seem to have evolved from entrapped bacteria.
Other examples include the presence of introns, and poly-A tails on mRNA.
This genus includes B. burgdorferi, the causative agent of Lyme disease.
Streptomyces make a wide variety of useful antibiotics, including streptomycin.
For example, linear DNA was precipitated in the most commonly used procedures for purifying bacterial plasmids, and the procedures for purifying chromosomal DNA relied upon the differential binding of ethidium bromide to "sheared DNA fragments" compared to circular DNA.
It is not intuitively obvious how the ends of a linear DNA molecule could be completely replicated. All known DNA polymerases require a pre-existing primer for initiation of DNA replication. The primer is usually a short RNA molecule with a free 3'-OH group that can be extended by DNA polymerase. If a linear DNA molecule was primed at one end, DNA synthesis could continue to the other end. However, once the primer is removed, the DNA corresponding to the primer could not be replicated.
The telomers at the end of chromosomes of most eukaryotic cells are replicated by a different mechanism: most telomeres are short GC-rich repeats that are added in a 5' to 3' direction by the enzyme telomerase (see Blackburn. 1991. Telomeres. Trends Biochem. Sci. 16: 378-381).
0 comments:
Post a Comment